芯弃疾JX-8B数字化高灵敏ELISA芯片检测产品;应用范围:各种高灵敏多重免疫检测,可替代各种ELISA试剂盒,及其他免疫检测产品。
参考原理:通过量化异常水平的生物标志物来检测新生疾病过程是诊断和治干预的关键,以在出现继发性临床症状之前进行干预。蛋白质和核酸生物标志物的发现和验证已成为生物制药研究、靶向临床研究设计以及早期疾病诊断追求的主要驱动力。1–4由于蛋白质生物标志物提供了比核酸更多的下游信息内容,因此它们可能作为临床决策工具具有比较大的潜力。5据估计,人类蛋白质组来源于超过20,000个基因,且循环中有超过4000种分泌蛋白。6,7这些分泌蛋白中不到十分之一(375种)可以通过蛋白质测定技术可靠地测量。6在这些可测量的蛋白质中,几乎有一半(171种)已由美国食品药品监督管理局(FDA)批准的诊断测试,8个指出了蛋白质生物标志物对人类健康的重要性。POCT 芯片搭配小型化设备,即时检测 hs-cTnT 等指标,为急诊救治提供快速数据支持。阵列单分子数字ELISA精密度

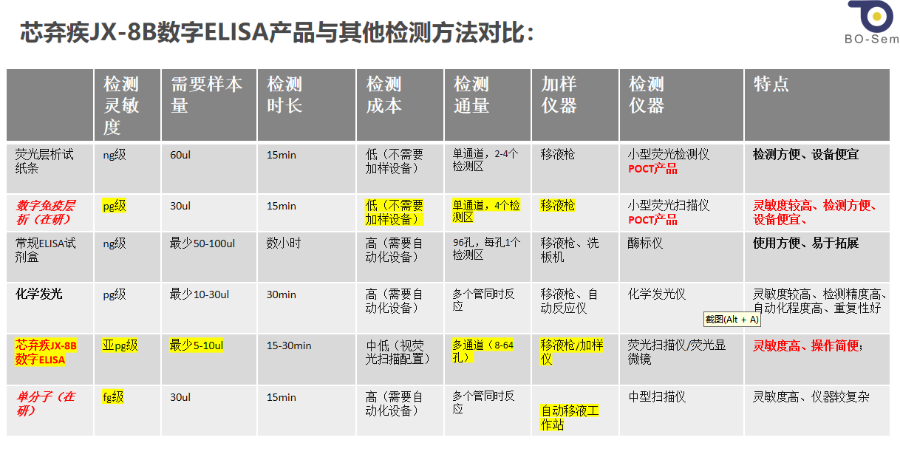
芯弃疾JX-8B数字ELISA高敏检测产品;具有以下特点:多重、超敏微量、极速灵活、开放;
只有少量分泌蛋白可测量的可能性突显了蛋白质测量领域面临的挑战:医学上相关的生物标志物可能存在于非常低的丰度中。免疫测定仍然是是蛋白质生物标志物敏感和特异性测量的基础。然而,传统的免疫分析技术在检测不可测量的生物标志物时灵敏度不足,这些生物标志物肯定位于当前可检测范围之下。主流的传统免疫分析方法——包括酶联免疫吸附试验(ELISA)、化学发光和电化学发光——的灵敏度下限约为10^-13M(~<0.1pM)。许多降低灵敏度的方法已被描述,包括拉曼增强信号检测、电感耦合等离子体质谱,但这些方法的数据表明其成功有限。非常规方法如亚飞摩尔级检测具有明显的权衡,例如程序较长或无法提供定量答案。
芯弃疾产品数字ELISA购买灵活抗体筛选芯片支持同一反应体系交叉测试,适合婴幼儿等困难场景。

芯弃疾JX-8B数字ELISA产品
每个生物实验室都用得起的单分子免疫检测
数字ELISA测量蛋白质浓度远低于传统ELISA的能力源于两种效应:1)SiMoA对酶标记的高度敏感性;以及2)通过数字化蛋白质检测可以实现的低背景信号。任何免疫测定的灵敏度由其灵敏度决定。检测技术到标签,抗体亲和力,试验背景,以及背景测量值的变异(%CV)27.SiMoA对酶非常敏感标签(图2)为在数字ELISA中检测亚飞摩尔浓度的标记蛋白提供了基础。也就是说,对于给定亲和力的抗体,其灵敏度为免疫测定将由测定背景决定,SiMoA的高标记灵敏度有助于降低这种背景。对照实验表明,数字ELISA的背景来自于检测抗体和酶的非特异性结合(NSB)与捕获珠表面结合(补充表2)。AsSiMoA相比传统检测方法具有更高的标记灵敏度,明显减少了检测抗体(~1nM)和酶标记物(1–50pM)的需要,以检测结合事件,与传统方法相比(标记试剂浓度~10nM)。降低的标记物浓度减少了NSB到捕获表面,从而导致背景信号明显降低。
芯弃疾JX-8B数字ELISA,我们为什么能做到?产品主要原理同单分子阵列技术:
非常近,已经描述了两种数字蛋白质测量方法,这些方法能够提高对单分子水平的灵敏度。一种方法依赖于在固相上形成免疫三明治复合物,然后化学解离并通过激光计数每个分子。第二种方法由美国开发,依赖于单分子阵列和同时计数单分子捕获微珠。这两种方法都能将检测能力的下限降低10倍或更多,与增强的模拟放大方法相比,但后者技术也易于与高通量自动化仪器兼容,用于ELISA试剂处理。通过使用大量微孔阵列,可以同时获取和查询数百到数万个数据点,实现快速数据采集和稳健统计。此外,从阵列中可能获得的快速数据采集可以应用于预编码具有不同荧光特性的多个微珠亚群,从而在单分子水平上实现高通量多重分析。 全自动加样与图像分析系统实现检测流程自动化,荧光信号识别,结果可靠。

芯弃疾JX-8B数字ELISA产品
每个生物实验室都用得起的单分子免疫检测
珠子以两种不同的方式读出。首先,在与100μMresorufin-阝-D孵育1小时后,用荧光板读出器以100μL方式读出珠子结合-半乳糖苷(RGP),一种荧光底物for阝-半乳糖苷酶。在平板阅读器上,检测限为15fMofS阝G(图2)。其次,将珠子加载然后将RGP溶液密封到阵列的孔中,单个酶的信号被允许在反应室中积累,并每30秒获取一次荧光图像。实验结束时获取阵列的白光图像以识别含有珠子的孔(含有珠子的孔散射光与空井不同)。荧光图像用于确定哪些微球具有相关的结合酶(从时间变化的荧光图像中强度增加)。图2显示了微球中含酶的比例与总体S阝G浓度的关系的对数-对数图。检测到的比较低酶偶联物浓度为350飞摩尔(zM),并通过外推信号等于的酶浓度计算得出的检测限(LOD)。 芯弃疾JX-8B简易版单分子ELISA检测产品, 极速检测,快至15min能完成 的ELISA检测!单分子级别数字ELISA开放
芯弃疾JX-8B单分子普惠化ELISA检测产品,超敏检测,理论可达飞克级;阵列单分子数字ELISA精密度
芯弃疾JX-8B数字ELISA产品
每个生物实验室都用得起的单分子免疫检测
通过SiMoA对酶标记物进行数字检测的线性动态范围由区分“开启”和“关闭”孔的能力决定。在酶与珠子的比例较低(小于约1:10)时,泊松统计表明,只有统计学上有效果的群体珠子是指含有零和一个酶的珠子。只要足够多的珠子被检测,单个酶就可以被检测到,并且活性珠子的数量会超过泊松分布计数活性微球的噪声。在酶与微球的高比率(大于约(1:10),活性珠子的比例变得更高,泊松统计表明有大量含有多种酶的微球。为了定量检测到的酶的数量并保持含有多种酶的微球亚群中的线性对于酶,我们使用泊松统计法将活性珠子的数量转换为检测到的酶的数量 阵列单分子数字ELISA精密度